研究报告/Research Report
黄连木PcSAD基因的克隆及分析 
章文乐  ,
蒲婧懿 ,
朱梦媛 ,
钮俊 ,
冯清茗 ,
胡惠雯 ,
侯琦 ,
郭静 ,
袁一鸣 ,
张志翔 ,
林善枝
,
蒲婧懿 ,
朱梦媛 ,
钮俊 ,
冯清茗 ,
胡惠雯 ,
侯琦 ,
郭静 ,
袁一鸣 ,
张志翔 ,
林善枝 
北京林业大学生物科学与技术学院, 林木育种国家工程实验室, 林木花卉遗传育种教育部重点实验室, 北京, 100083
作者 通讯作者
植物药与药理学杂志, 2013 年, 第 2 卷, 第 1 篇 doi: 10.3969/gab.032.000503
收稿日期: 2013年08月28日 接受日期: 2013年08月30日 发表日期: 2013年09月07日
作者 通讯作者
植物药与药理学杂志, 2013 年, 第 2 卷, 第 1 篇 doi: 10.3969/gab.032.000503
收稿日期: 2013年08月28日 接受日期: 2013年08月30日 发表日期: 2013年09月07日
© 2013 BioPublisher 生命科学中文期刊出版平台
本文首次发表在 《基因组学与应用生物学》上。现依据版权所有人授权的许可协议,采用 Creative Commons Attribution License,协议对其进行授权,再次发表与传播。只要对原作有恰当的引用, 版权所有人允许并同意第三方无条件的使用与传播。
推荐引用:
引用格式(中文):
章文乐等, 2013, 黄连木PcSAD基因的克隆及分析, 植物药与药理学杂志(online) Vol.2 No.1 pp.1-7 (doi: 10.3969/gab.032.000503)
引用格式(英文):
Zhang W.L., et al., 2013, Cloning and analysis of Pc SAD Gene from Pistacia chinensis Bunge, Zhiwuyao Yu Yaolixue Zazhi (online) Vol.2 No.1 pp.1-7 (doi: 10.3969/gab.032.000503)
摘要
中国黄连木(Pistacia chinensis Bunge)是种重要的优良木本油料树种,分布广,而且种子具有含油量高、出油率高、油品质好等优点,是生产生物柴油的理想原料。硬脂酰 - 酰基载体蛋白脱饱和酶(stearoyl-ACP desat-urase, SAD)在植物中催化不饱和脂肪酸生物合成的第一步脱饱和反应,对植物细胞膜和种子油脂中脂肪酸成分的调控起着重要作用。本研究采用 RACE 及 RT-PCR 方法,首次从黄连木种子中克隆获得长度为 1 791 bp的 SAD 基因(命名为 Pc SAD),该基因含有 1 299 bp 完整的开放阅读框、编码 432 个氨基酸。生物信息学分析发现,黄连木 Pc SAD 基因的核苷酸序列及其编码的氨基酸序列与已报道的其它相近物种具有较高同源性,而且 Pc SAD 含有典型的酰基载体蛋白脱饱和酶 2 和类铁蛋白家族的保守结构域;另外,开展了 Pc SAD 的原核表达载体构建及诱导表达体研究。上述研究结果可为黄连木 Pc SAD 基因的功能鉴及应用定奠定重要基础。
关键词
黄连木;硬脂酰 - 酰基载体蛋白脱饱和酶;基因克隆;原核表达;生物信息学分析
随着一次化石能源危机加剧,清洁型可再生性生物质能源的开发和高效利用已成为人们研究热点。全球植物每年贮存的能量约相当于世界主要燃料消耗的10倍(蒋建春和应浩, 2005),从含有大量油脂的植物种子中提取液体燃油、直接用于燃烧或将其酯化处理后制取可替代一次能源柴油的生物质燃油的方法,是近年来生物质燃料研究的重点。目前,美、英、法等国均已建立了不少生物柴油公司,但其原料多以油料农作物为主,这对人类食物供应保障提出了挑战;而木本植物具有物种丰富、能量密度高、一次种植且多年受益等优点(蒋建新等, 2005),备受研究者关注。因此,开展木本油料生物质能源研究,对减少石化燃料的需求、缓解人类所面临的资源短缺及环境污染等诸多方面具有十分重要意义。
植物硬脂酰酰基载体蛋白脱饱和酶(stearoyl-acylcarrier protein desaturase, SAD)是一种可溶性质体酶,可催化硬脂酰 -ACP (18:0-ACP)在其中介入第 1 个双键而生成油酰基 -ACP (18:1-ACP) (Thompson et al., 1991; Ohlrogg and Browse, 1995),进而在其它酶的作用下形成甘油酯或多不饱和脂肪酸。由此可见,SAD 催化的脱饱和作用是植物脂肪酸代谢途径中不饱和脂肪酸合成的关键步骤,起着调控植物贮脂中饱和脂肪酸与不饱和脂肪酸比例的作用。Shanklin和Somerville (1991) 研究发现,硬脂酰-ACP m RNA 在发育的种子中的含量远高于叶片和根部组织,这表明脱饱和酶的表达是在发育中受调控的。至今已有研究将植物 SAD 基因的反义抑制 (Kuntzon et al.,1992)、RNA 干扰(Liu et al., 2002)、超表达和核酶(Merlo et al., 1998)等基因工程技术应用于农作物,且均在转基因作物中成功检测到 C16及 C18的饱和与不饱和脂肪酸在含量和组成上的改变。尽管目前已从红花、蓖麻、胡麻、黄瓜、油菜、胡麻、菠菜等草本植物分离获得 SAD 基因,但木本植物的相关研究却相对较少。
黄连木(Pistacia chinensis Bunge)为漆树科(Anac-ardiaceae)黄连木属(Pistacia)落叶乔木,在我国分布广泛,是一种优良的绿化、用材、观赏、药用和油料树种(赵和文等, 2004);而且黄连木种子含油量49.3%,种子油脂中脂肪酸组成主要为棕榈酸、亚油酸和油酸,其中主要不饱和脂肪酸含量高达 82.4% (蒲志鹏等, 2009)。另外,生物柴油提取工艺的研究表明,黄连木种子出油率高、油品好,所生产的生物柴油的碳链长度集中在 C17~C20之间,与普通柴油主要成分的碳链长度极为接近(侯新村, 2006)。据此,本研究以黄连木近成熟种子为试材,通过 RT-PCR 和 RACE 方法克隆 Pc SAD 基因的 c DNA 序列,并分析其编码蛋白质的空间结构和功能位点,为进一步研究 Pc SAD基因功能及 Pc SAD 调节不饱和脂肪酸含量机制奠定重要基础。
1结果与分析
1.1黄连木种子 Pc SAD 基因的克隆与分析
从黄连木种子 c DNA 中扩增得到一个 420 bp 的条带(图 1A),测序结果显示该片段与 Gen Bank 上 EF-179606 中提供的黄连木 c DNA 序列一致。通过 3'RACE 和5' RACE 的扩增延伸,分别得到长度为 1 259 bp 和984 bp 的两个片段(图 1 B; 图 1 C),三个片段去重拼接后得到全长为 1 791 bp 的 Pc SAD cDNA。序列分析结果表明,该 cDNA 序列含有一个 1 299 bp 的完整开放阅读框(ORF) (图1 D),其编码的蛋白含 432 个氨基酸残基,相对分子量为49.1 kD,等电点为6.46,含有42.6%疏水氨基酸、29.6%亲水氨基酸、14.6%碱性和13.2%酸性氨基酸(图2)。
|
图 1 黄连木 Pc SAD 基因的克隆
注: A: 已知的 Pc SAD 基因 m RNA 中间片段的扩增; B: 3'RACE 基因下游序列的扩增; C: 5' RACE 基因上游序列的扩增; D: Pc SAD 基因开放阅读框 c DNA 全长序列的扩增
Figure 1 Cloning of Pc SAD gene from Pistacia chinensis
Note: A: Amplification of the known segment of Pc SAD; B: Am-plification of the down-stream segment (3' RACE); C: Amplification of the up-stream segment (5' RACE); D: Amplification of Pc SAD ORF
|
.png)
|
图 2 黄连木 Pc SAD 基因的 c DNA 序列及其推导的氨基酸序列
注: 图中用“→”标注出编码的 Pc SAD 基因的开放阅读框
Figure 2 Nucleotide and deduced amino sequences of the full-length c DNA sequence of Pc SAD gene from P. chinensis
Note: The ORF of Pc SAD was maked by“→”
|
1.2 PcSAD 基因序列比对和 SAD 系统进化树分析
黄连木 PcSAD 基因与其它物种 SAD 基因的核苷酸序列和氨基酸序列比对分析表明,黄连木 PcSAD 基因与蓖麻,木油桐等油料植物 SAD 基因均具有较高的同源性,尤其与蓖麻 SAD 核苷酸序列和氨基酸序列的同源性分别高达 86%和 91% (表 1)。
|
表 1 黄连木与其它物种 SAD 基因的核苷酸序列和氨基酸序列的一致性
Table 1 Identities of nucleotides and peptides of SADs between P. chinensis and other plants
*注: N: 核苷酸; P: 蛋白质
Note: N: Nucleic acid; P: Protein
|

对包括黄连木在内的共 17 种植物的 SAD 构建发育进化关系树,由图 3 可知,进化树可分为四个支系,第一分支有 10 种 SAD,属于六个科的植物;第二分支有 7 种 SAD,属于四个科的植物;第三分支只有1 种 SAD;第四分支有 3 种 SAD,属于两个科的物种。而黄连木与蓖麻、麻疯树、油桐、胡麻等油料植物同在第一个分支。
|
图 3 17 种植物 SAD 的 N-J 系统进化树分析
注: 17 种植物包括: 木油桐; 油桐; 乌桕; 麻疯树; 蓖麻; 胡麻;毛白杨; 黄连木; 葡萄; 棉豆; 蒺藜苜蓿; 百脉根; 大豆; 籼稻; 眼黑花; 阿甘树; 橄榄
Figure 3 Phylogenetic N-J tree analysis of SADs from 17 plants
Note: 17 plant species: Vernicia Montana; Vernicia fordi; Triadica sebifera; Jatropha curcas; Ricinus communis; Sesamum indicum; Populus trichocarpa; Pistacia chinensis; Vitis viniferagi; Phaseolus lunatus; Medicago truncatula; Lotus japonicas; Glycine max; Oryza sativa; Thunbergia alata; Argania spinosa; Olea europaea
|

1.3 Pc SAD 基因的原核表达及其编码蛋白的结构预测
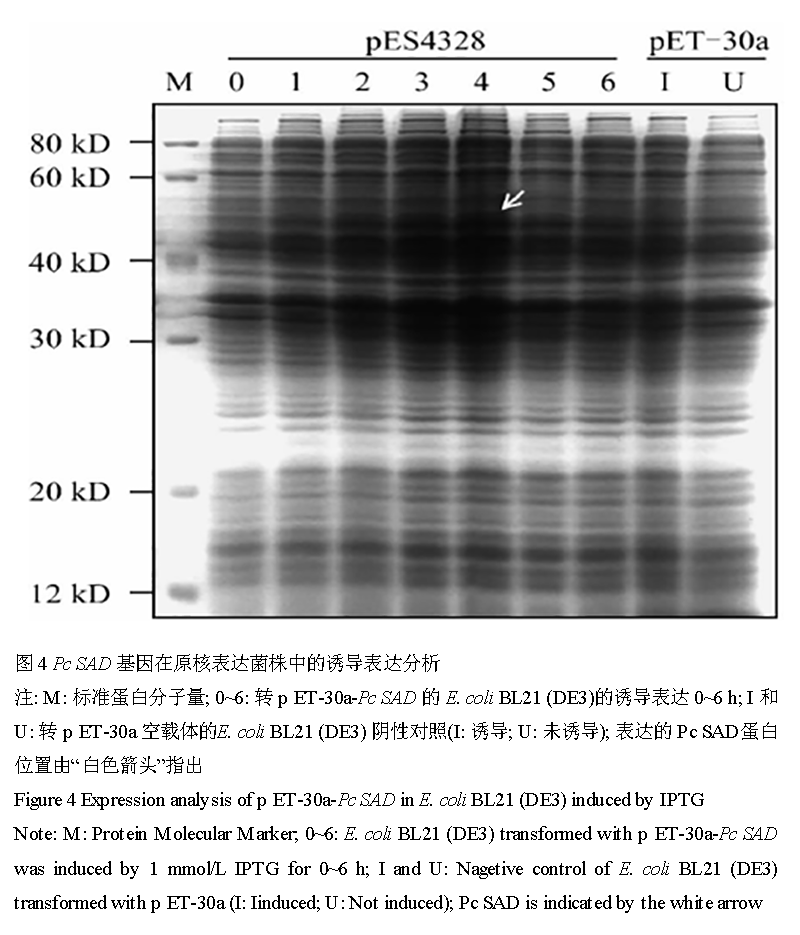
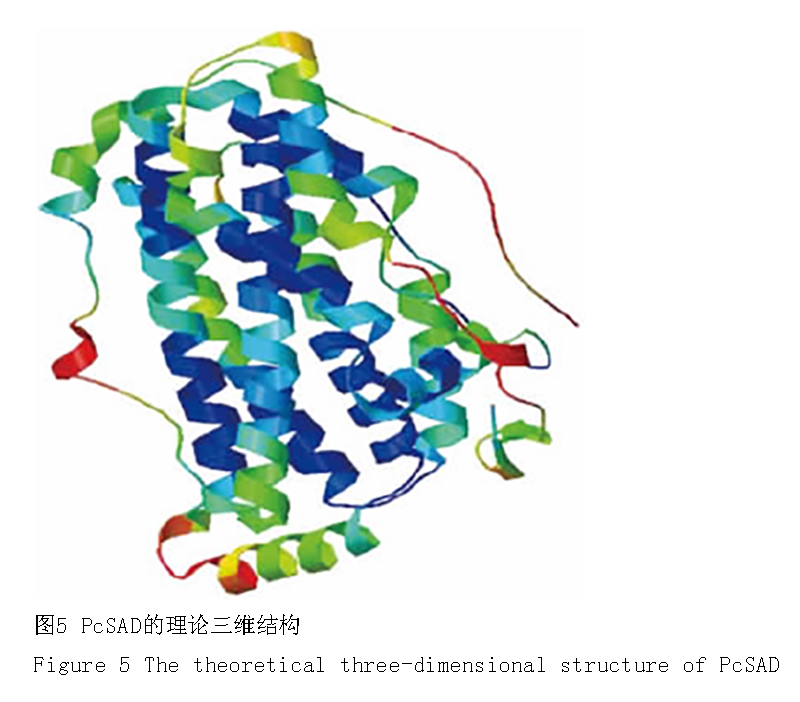
原核诱导表达后收集的菌液经 SDS-PAGE 电泳分析后发现,重组质粒 p ES4328 经 IPTG 诱导 4 h 后可表达得到含量较高的分子量约 50 k D 的特异性蛋白;而阴性对照中并无明显被诱导表达的蛋白出现(图 4)。Pc SAD 蛋白氨基酸序列的保守结构域分析后发现,Pc SAD 具有酰基载体蛋白脱饱和酶 2 (Acyl-ACPDesaturase_2)家族和类铁蛋白(ferritin-like)家族的高保守结构域;而且在 Pc SAD 与 Acyl-ACP Desat-urase_2 家族和 ferritin-like 家族对应的高保守区重叠的氨基酸残基序列中,包含植物 SAD 活性相关的6 个 diiron 中心结合位点、33 个二聚体表面结合位点和 14 个底物结合口袋形成位点(Fox et al., 1994)。另外,Pc SAD 空间构象分析显示,其二级结构主要由11 个 α- 螺旋、2 个 β- 折叠和大部分的无规则卷曲组成(图 5),由此推断 Pc SAD 立体模型与蓖麻的晶体结构(Lindqvist et al., 1996) 极为相似。
|
图 4 Pc SAD 基因在原核表达菌株中的诱导表达分析
注: M: 标准蛋白分子量; 0~6: 转 p ET-30a-Pc SAD 的 E. coli BL21 (DE3)的诱导表达 0~6 h; I 和 U: 转 p ET-30a 空载体的E. coli BL21 (DE3) 阴性对照(I: 诱导; U: 未诱导); 表达的 Pc SAD蛋白位置由“白色箭头”指出
Figure 4 Expression analysis of p ET-30a-Pc SAD in E. coli BL21 (DE3) induced by IPTG
Note: M: Protein Molecular Marker; 0~6: E. coli BL21 (DE3) transformed with p ET-30a-Pc SAD was induced by 1 mmol/L IPTG for 0~6 h; I and U: Nagetive control of E. coli BL21 (DE3) transformed with p ET-30a (I: Iinduced; U: Not induced); Pc SAD is indicated by the white arrow
|
|
图 5 图5 PcSAD的理论三维结构
Figure 5 The theoretical three-dimensional structure of PcSAD
|
2讨论
本研究采用异源的原核表达系统 E. coli BL21(DE3)作为 Pc SAD 重组体的宿主菌,其优点为 BL21作为野生型大肠杆菌,以厌氧途径合成不饱和脂肪酸,自身不存在脂肪酸脱饱和酶,可排除实验干扰。脂肪酸脱饱和酶需要依赖铁氧还蛋白(Cahoon et al., 1996)或者细胞色素 b5 (Petrini et al., 2004)提供电子才能具有活性,造成许多在 E. coli 中异源表达的脱饱和酶为失活状态,无法体现其体内功能;而黄连木Pc SAD 为可溶性质体蛋白并非膜整合蛋白,E. coli虽不具有植物 SAD 理想电子供体铁氧还蛋白和铁氧还蛋白-NADP+氧化还原酶系统,但其胞质中存在的细胞色素 b5 可提供植物 SAD 所需的电子,并不影响植物 SAD 在其中顺利表达(Cao et al., 2010)。
许多研究发现,植物体中的 SAD 大多是以同源二聚体形式存在,最早证实 SAD 的二聚体天然形态是从红花中分离纯化得到大约 200 倍纯度的红花SAD,其分子量为 68 kD (Mckeon and Stumpf, 1982)。SAD 的二聚体中的每个单体都有独立的 diiron-oxo活性结合位点,位于二聚体分子表面的凹槽内部,特异性的底物硬脂酰 -ACP 分子通过嵌入与位于凹槽底部的活性中心结合,进而发生脂肪酸碳链 C9~C10的氧化还原反应(Lindqvist et al., 1996; Browse, 1996)。在本研究中,通过对黄连木 Pc SAD 编码的氨基酸序列比对分析后发现,Pc SAD 具有很高的保守性,推测其可能在黄连木中同样以同源二聚体形式存在,并含有二聚体表面结合位点和酶活性位点,可对硬脂酰-ACP 行使催化活性;另外,通过 Pc SAD 基因的原核诱导表达研究,成功获得分子量约为 50 k D 的蛋白质,推测该蛋白可能是黄连木 Pc SAD 天然二聚体的寡聚单体形态。
Harwood (1998)指出植物脂肪酸合成途径中的脱饱和酶主要富集于前质体或叶绿体基质中,而且从红花(Thompson, 1991)、蓖麻(Fox et al., 1994)和麻疯树(Luo et al., 2007)等植物中克隆得到的 SAD 氨基酸序列的 N 端均含有转运肽。但我们前期研究发现,黄连木 Pc SAD 氨基酸序列的信号肽分析结果显示其 N 端不存在信号肽,而且通过亚细胞定位预测分析后发现 Pc SAD 氨基酸序列 N 端含有叶绿体导肽裂解位点,这说明黄连木 Pc SAD 基因编码的可能是一个定位于叶绿体内的硬脂酰 -ACP 脱饱和酶。
3材料与方法
3.1实验材料
黄连木种子取自河北省唐县,采集后用液氮速冻并保存于 -70℃冰箱。选取 3~6 颗铜绿色黄连木种子,置于预冷过的无 RNase 研钵中用液氮浸泡研磨成适当粗细的粉末(Andrew and Brian, 1976),提取RNA直接使用或保存于 -80℃。
3.2黄连木种子总RNA 的提取及反转录
黄连木种子总 RNA 采用 RNeasy Plant Mini Kit(QIAGEN)试剂盒提取。c DNA 第一链反转录合成采用 Plexor R One-Step q RT-PCR System (Promega)反转录试剂盒。根据 Gen Bank 中已知的黄连木保守硬脂酰酰基载体蛋白脱饱和酶(EF179606.1) m RNA 片段的保守序列,设计上、下游引物(上游引物 5'-AGCCCTTCCTACTTATCAAACAA-3', 下 游 引 物 5'-CATCAGCAGCAATTATACCACAT-3'),以黄连木种子 c DNA 为模板进行 PCR 扩增。
3.3 Pc SAD 基因c DNA 的克隆
采用 3'-Full RACE Core Set Ver.2.0 (Ta Ka Ra)和 SMARTer TM RACE c DNA amplification Kit (Clon-tech)试剂盒,以总 RNA 为模板,分别获得黄连木种子的 3' RACE cDNA 和 5' RACE cDNA。根据已知SAD cDNA 序列设计 3' 和 5' RACE 所需的基因特异性引物。以 3' RACE cDNA 为模板,利用 3' 末端外侧特异引物(5'-ATCAGCAGCAATTATACCACATT-3')与试剂盒 3' RACE 外侧引物(5'-TACCGTCGTTCCACTAGTGATTT-3')作首轮 PCR 扩增;采用 3' 末端内侧特异引物(5'-CCCTTCCTACTTATCAAACAATGC-3')与试剂盒 3' RACE 内侧引物(5'-CGCGGATCCTCCACTAGTGATTTCACTATAGG-3')进行巢式 PCR。以5' RACE c DNA 为模板,利用 5' 末端特异引物(5'-CAGTGGTATCAACGCAGAGT-3') 与试剂盒 5' RACE外侧短引物(5'-CAGTGGTATCAACGCAGAGT-3')进行 PCR 扩增。
将 3' RACE 和 5' RACE 扩增产物测序后拼接获得完整的 SAD 基因序列,根据 NCBI 分析所得 ORF设计 SAD 基因全长引物(上游引物: 5'-ATGGGGATTGTTCAACTCACTTT-3'; 下游引物: 5'-TTAAAGCTGTACTTGTCTGTCGAAA-3'),以黄连木种子 c D-NA 为模板进行 PCR 扩增。引物合成和测序分别由上海生工和金唯智生物科技(北京)有限公司完成。
3.4原核表达载体构建和诱导表达
在 Pc SAD 两端通过 PCR 增加 Xba Ⅰ和 Sac Ⅰ酶切位点,并回收 PCR 产物;经测序鉴定后,在 T4 连接酶作用下,与经 Xba Ⅰ和 SacⅠ双酶切后回收的p ET-30a(+)载体链接,构建含有 Pc SAD 基因特异序列的重组质粒 p ES4328,转化至原核表达系统 E. coli BL21 (DE3)。从平板上挑取阳性重组表达单菌落于 2 m L 含抗生素的 LB 培养基内,在 37℃、200 r/min 下培养过夜;按 1:50 的接种量转种于含抗生素的营养丰富型培养基 2×YT (胰蛋白胨 16 g, 酵母提取物 10 g, NaCl 5 g) 中,在 37℃、200 r/min 下培养至 OD600=0.4~0.6,加入终浓度为 0.1 mol/L 的 IPTG,25℃诱导表达 6 h,每隔 1 h 取样 1 次;4℃、8 000×g 离心 5 min 收集菌体,等量稀释后与 2×SDS-PAGE 上样缓冲液(100 mmol/L Tris-Cl p H 6.8, 4% SDS、200 mmol/L DTT 或者巯基乙醇, 0.2%溴酚蓝, 20%甘油)等体积混匀,100℃水浴5 min,12 000×g 离心 1 min,取其上清进行 SDS-PAGE(12%分离胶)电泳分析,考马斯亮蓝 R-250 染色。
3.5 Pc SAD 基因编码蛋白的生物信息学分析
利用 NCBI 数据库搜索相关植物的 SAD 蛋白及其编码基因(Altschul et al., 1997),并在保守结构域数据库(CDD)中进行序列比对(Marchler and Bryant, 2004);采用 DNA Tool 5.1 软件分析 Pc SAD 氨基酸序列;运用 Ex PASY Proteomics Server (http://cn.ex-pasy.org/) 和 Swiss-Model (http://swissmodel.expasy.org/)分析蛋白的结构域、功能位点和空间构象;利用Clustal X 2.0 进行多序列比对分析,运用 MEGA3 软件构建系统进化树。
作者贡献
通讯作者林善枝负责实验指导及论文修改;章文乐和蒲婧懿为共同第一作者,负责整个实验的具体实施及论文撰写;钮俊和侯琦负责基因克隆所需引物的设计及部分数据分析;胡惠雯和朱梦媛负责种源收集等;其他作者均参与了本实验的具体实施工作。
致谢
本项目由林业公益性行业科研专项(201004001)和“十二五”农村领域国家科技计划课题(2011BAD22B08)共同资助。
Altschul S.F., Madden T.L., Schaffer A.A., Zhang J., Zhang Z., Miller W., and Lipman D.J., 1997, Gapped BLAST and PSI-BLAST: A new generation of protein database search programs, Nucleic Acids Research, 25: 3389-3402
Andrew O.J., and Brian A.L., 1976, Influence of ionic strength, p H, and chelation of divalent metals on isolation of polyribosomes from tobacco leaves, Plant Physiology, 57: 5-10
Browse J., 1996, Towards rational engineering of plant oils: Crystal structure of the 18:0-ACP desaturase, Trends in Plant Science, 1: 403-404
Cahoon E.B., Mills L.A., and Shanklin J., 1996, Modification of the fatty acid composition of Escherichia coli by coexpression of a plant acyl-acyl carrier protein desaturase and ferredoxin, Journal of Bacteriology, 178: 936-939
Cao Y.J., Xian M., Yang J.M., Xu X., Liu W., and Li Z.L., 2010, Heterologous expression of stearoyl-acyl carrier protein desaturase (S-ACP-DES) from Arabidopsis thaliana in Escherichia coli, Protein Expression and Purification, 69: 209-214
Fox B.G., Shanklin J., Ai J.Y., Loehr T.M., and Sanders L.J., 1994, Resonance raman evidence for an Fe-O-Fe center in stearoyl-ACP desaturase, primary sequence identify with other Diiron-Oxo proteins, Biochemistry, 33: 12776-12786
Fox B.G., Shanklin J., Somerville C., and Munck E., 1993, Stearoylacyl carrier protein △9 desaturase from Ricinus communis is a diiron-oxo protein, PNAS, 90: 2486-2490
Harwood J.L., 1988, Fatty acid metabolism, Annual Review Plant Physiology and Plant Molecular Biology, 39: 101-138
Hou X.C., 2006, Investigation and research on woody energy plant Pistacia chinensis for biodiesel, Thesis for M.S., Chinese Academy of Forestry, Supervisors: Wang T., and Gao C.M., pp.89-90 (侯新村, 2006, 生物柴油木本能源植物中国黄连木(Pistacia chinensis)的调查与研究, 硕士学位论文, 中国林业科学研究院, 导师: 王涛, 高崇明, pp.89-90)
Jiang J.C., and Ying H., 2005, The trand of energy conversion technology of forestry biomass in China, Linchan Huaxue Yu Gongye (Chemistry and Industry of Forest Products), 25: 5-9 (蒋建春, 应浩, 2005, 中国林业生物质能源转化技术产业化趋势, 林产化学与工业, 25: 5-9)
Lindqvist Y., Huang W., Schneider G., and Shanklin J., 1996, Crystal structure of △9 stearoyl-acyl carrier protein desaturase from castor seed and its relationship to other di-iron proteins, The EMBO Journal, 15(16): 4081-4092
Liu Q., Singh P.S., and Green G.A., 2002, High-stearic and High-oleic cottonseed oils produced by hairpin RNA-mediated post-transcriptional gene silencing, Plant Physiology, 129: 1732-1743
Luo T., Ma D.W., Xu Y., Deng W.Y., Xiao M., Qing R.W., and Chen F., 2007, Cloning and characterization of a stearoyl-ACP desaturase gene from Jatropha curcas, Journal of Shanghai University, 11(2): 182-188
Marchler B.A., and Bryant S.H., 2004, CD-Search: Protein domain annotations on the fly, Nucleic Acids Research, 32: 327-331
Mckeon T.A., and Stumpf P.K., 1982, Purification and characterization of the stearoyl-acyl carrier protein desaturase and the acyl-acyl carrier protein thioesterase from maturing seeds of safflower, Journal of Biological Chemistry, 257: 12141-12147
Merlo A.O., Cowen N., Delate T., Edington B., Folkerts O., Hopkins N., Lemeiux C., Skokut T., Smith K., Woosley A., Yang Y.J., Young S., and Zwickb M., 1998, Ribozymes targeted to stearoyl-ACP △9 desaturase m RNA produce heritable increases of stearic acid in transgenic maize leaves, The Plant Cell, 10: 1603-1621
Petrini G.A., Altabe S.G., and Uttaro A.D., 2004, Trypanosoma brucei oleate desaturase may use a cytochrome b5-like domain in another desaturase as an electron donor, European Journal of Biochemistry, 271: 1079-1086
Pu Z.P., Wang W.G., Jiang J.X., Yang T., and Ma L., 2009, Biodiesel derived from Pistacia chinensis and its low-temperature fluidity, Beijing Linye Daxue Xuebao (Journal of Beijing Forestry University), 31(Supp.1): 56-61 (蒲志鹏, 王伟刚, 蒋建新, 杨婷, 马蕾, 2009, 黄连木生物柴油及其低温流动性能研究, 北京林业大学学报, 31(Supp.1): 56-61)
Shanklin J., and Somerville C., 1991, Stearoyl-acyl-carrier-protein desaturase from higher plants is structurally unrelated to the animal and fungal homologs, PNAS, 88: 2510-2514
Thompson G.A., Scherer D.E., Foxall-Van Aken S., Ke Nly J.W., Young H.L., Shintani D.K., Kridl J.C., and Knauf V.C., 1991, Primary structures of the precursor and mature forms of stearoyl-acyl canier protein desaturase from safflowerembryos and requirement of ferredoxin for enzyme activity, PNAS, 88: 2578-2582
Zhao H.W., Liu Z.L., Yu J.J., and Yang N.P., 2004, Studies on seed treatment of Pistacia chinensis Bunge (in Chinese), Beijing Nongxueyuan Xuebao (Journal of Beijing Agricultural Col-lege), 19(3): 42-45 (赵和文, 柳振亮, 于建军, 杨宁平, 2004, 黄连木种子处理的研究, 北京农学院学报, 19(3): 42-45)